Биодоступность железа и влияние ионов кальция на ее эффективность
Содержание железа в организме человека составляет в среднем 4,2 г. Около 75% от его общего количества входит в состав гемоглобина эритроцитов, которые переносят кислород от легких к тканям, 20% железа является резервным (костный мозг, печень, макрофаги), 4% входит в состав миоглобина, около 1% содержится в дыхательных ферментах, катализирующих процессы дыхания в клетках и тканях, а также в других ферментативных структурах. Железо осуществляет свою биологическую функцию, находясь в составе биологически активных соединений, преимущественно ферментов. Железосодержащие ферменты выполняют следующие основные функции:
- транспорт электронов (цитохромы);
- транспорт и депонирование кислорода (гемоглобин, миоглобин);
- участие в формировании активных центров;
- окислительно-восстановительные функции (оксидазы, гидроксилазы, супероксиддисмутазы и др.);
- транспорт и депонирование железа в плазме крови (трансферрин, ферритин).
Железо обладает несколькими особыми свойствами, которые отличают его от других биологически активных ионов и веществ.
В организме человека нет никаких специальных механизмов для выведения железа. В основном железо выделяется через кожу и кишечник (I. Guinote et al., 2006). Кроме этого, оно теряется также с волосами, ногтями, мочой и потом. Общее количество выделяемого железа у здорового человека (мужчины) составляет около 1 мг в сутки. Такое же количество в норме усваивается из потребляемой пищи (Linder, 1991). Отличие составляет менструальный период, когда потребление должно составлять около 4 мг железа в день. Таким образом, концентрация элемента в сыворотке крови зависит от его всасывания в желудочно-кишечном тракте, от накопления в селезенке, костном мозге и скелетных мышцах (миоглобин), а также от синтеза и распада гемоглобина и выделения его из организма. В пище железо может присутствовать в двух видах — гемовое и негемовое, которые характеризуются разными механизмами всасывания. Гемовое железо (порфириновое кольцо с атомом железа в центре, связанное с 4 атомами азота) в желудочно-кишечном тракте освобождается от белковых цепей и в виде металлопорфирина всасывается энтероцитами кишечника. Там происходит неспецифическое эндосомальное проникновение гема в клетку с последующим его разрушением. Далее, с помощью белковой транспортной системы IREG1, ионы железа окисляются до трехвалентного железа, связываются с трансферрином и покидают энтероцит, выходя в ток крови (Linder et al., 2006). В плазме крови железо перемещается в соединении с этим же белком, который выполняет как функцию депо, так и функцию переносчика. Наличие свободных ионов железа в крови не характерно и является патологией. Всасывание гемового железа происходит в пределах 15–50% (в среднем 20–30%).
Негемовое двухвалентное железо в желудке связывается белком гастроферрином и транспортируется в кишечник. Попадая в двенадцатиперстную кишку и проксимальную часть тощей кишки, железо проникает в энтероцит с помощью неспецифического ионного транспортера DMT1 (Divalent metal transport). Этот протонзависимый переносчик также участвует в транспорте многих других двухвалентных катионов, таких как Mn, Сu и Zn (M. Arredondo еt al., 2006). Кроме того, показано, что этот переносчик может транспортировать и некоторые одновалентные ионы, такие как Cu+1, который образуется при действии аскорбата на Cu+2 (M. Linder et al., 2006). Таким образом, можно предположить, что в зависимости от концентрации этих ионов в диете или мультивитаминной таблетке возможна их конкуренция за транспортер DMT1. При этом имеются данные о том, что существует специфичность переносчика DMT1 по отношению к разным двухвалентным ионам, связанная с их расположением по ходу желудочно-кишечного тракта, что было показано путем транскрипции различных железонезависимых mRNA транспортера DMT1, и конкуренции за переносчик не существует.
В литературе описывается несколько механизмов транспортировки железа внутри энтероцита, основанные главным образом на экспериментах, проведенных на культуре ткани Caco2 (M. Linder et al., 2006). Согласно первой теории, двухвалентное железо, поступившее в энтероцит с помощью транспортера DMT1, доставляется посредством везикул с трансферрином (некоторые отводят ему роль внутриклеточного рецептора) или в свободном состоянии к базолатеральной мембране энтероцита, где присутствует другой транспортер — IREGI/ferroportin/MTP1 (Donovan et al., 2000). Этот транспортер окисляет двухвалентное железо до трехвалентного и транспортирует в кровь, где он соединяется с плазменным трансферрином. Согласно второй теории, внутри энтероцита железо транспортируется, по-видимому, в везикулах вместе с апотрансферрином, который путем эндоцитоза попадает из тока крови в энтероцит (эндо-/экзоцитоз) (Ma et al., 2002). Во время этой транспортировки двухвалентное железо окисляется до трехвалентного и путем экзоцитоза поступает через базилярную мембрану энтероцита в кровь. В этом процессе возможно участие уже упоминавшейся системы IREG. Согласно литературным данным, именно механизм транспорта железа через базолатеральную мембрану энтероцита в кровь является лимитирующим в процессе адсорбции железа (Roy and Enns, 2000). Усвоение неорганического двухвалентного железа обычно происходит в пределах 6–15%.
Негемовое трехвалентное железо может быть восстановлено с помощью ферриредуктазы до двухвалентного железа и усвоено с помощью DMT1. Восстановление трехвалентного железа сильно зависит от кислотности желудочного сока. Невосстановленное железо может всасываться с помощью специфической интегрин-мобифериновой системы IMP. Усвоение трехвалентного железа происходит наименее полно и редко превосходит 4%.
Количество железа, поступающего в эффекторную клетку, куда оно транспортируется с кровью, прямо пропорционально числу мембранных рецепторов. В клетке происходит высвобождение железа из трансферрина. Затем плазменный апотрансферрин возвращается в циркуляцию. Повышение потребности клеток в железе при их быстром росте или синтезе гемоглобина ведет к индукции биосинтеза рецепторов трансферрина и, напротив, при повышении запасов железа в клетке число рецепторов на ее поверхности снижается. Железо, высвободившееся из трансферрина внутри клетки, связывается с ферритином, который доставляет микроэлемент в митохондрии, где он включается в состав гема. Помимо синтеза гема, двухвалентное железо используется в митохондриях для синтеза железосерных центров. В организме человека происходит постоянное перераспределение железа. В количественном отношении наибольшее значение имеет метаболический цикл: плазма — красный костный мозг — эритроциты — плазма. Обычно 70% плазменного железа поступает в костный мозг. За счет распада гемоглобина в сутки высвобождается около 21–24 мг железа, что во много раз превышает поступление железа из пищеварительного тракта (1–2 мг/сут).
Существует выраженная обратная зависимость между обеспеченностью организма железом и его всасыванием в пищеварительном тракте. В основном всасывание железа происходит в двенадцатиперстной кишке и проксимальных отделах тощей кишки и отсутствует в подвздошной кишке.
Всасывание железа зависит от следующих причин: возраста, обеспеченности организма железом, состояния желудочно-кишечного тракта, количества и химических форм поступающего железа и прочих компонентов пищи. Для оптимального всасывания железа необходима нормальная секреция желудочного сока. Прием соляной кислоты способствует усвоению железа при ахлоргидрии. В таблице приведены основные вещества, содержащиеся в продуктах питания человека, которые могут активизировать или уменьшать всасывание железа, содержащегося в этих продуктах или мультивитаминной таблетке. Аскорбиновая кислота, восстанавливающая железо и образующая с ним хелатные комплексы, повышает доступность этого элемента так же, как и другие органические кислоты. Она является одним из наиболее сильных стимуляторов всасывания железа. Другим компонентом пищи, повышающим всасывание железа, является «фактор животного белка», в котором содержится миоглобин и гемоглобин. Улучшают всасывание железа простые углеводы: лактоза, фруктоза, сорбит, а также такие аминокислоты, как гистидин, лизин, цистеин, образующие с железом легковсасываемые хелаты.
Самыми сильными ингибиторами, блокирующими всасывание железа, являются фитаты и полифенолы. Фитаты представляют собой форму хранения фосфатов и минералов, присутствующих в зернах злаковых растений, овощах, семенах и орехах. Они активно тормозят всасывание железа, действуя при этом в прямой зависимости от дозы. Всасывание железа снижают такие напитки, как чай, содержащий таннин, а также другие полифенольные соединения, которые прочно связывают этот элемент. Феноловые соединения существуют почти во всех растениях и являются частью системы защиты против насекомых и животных. Поэтому чай применяют для профилактики повышенного усвоения железа у больных талассемией. Большое влияние на усвоение железа оказывают различные заболевания. Оно усиливается при недостаточности железа, при анемиях (гемолитической, апластической, пернициозной), гиповитаминозе В6 и гемохроматозе, что объясняется повышением эритропоэза, истощением запасов железа и гипоксией.
Из перечисленных веществ, которые могут уменьшать всасывание железа, особое внимание обращает на себя ион кальция. Кальций обладает высокой биологической активностью, в значительном количестве содержится в основных продуктах питания и, как правило, присутствует в одной мультивитаминной таблетке с железом.
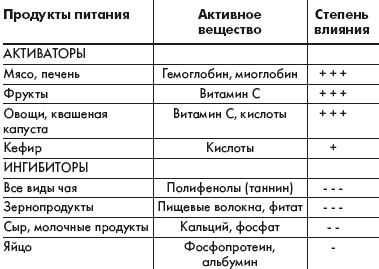
|
| Таблица. Активаторы и ингибиторы всасывания железа, содержащиеся в пищевом рационе человека |
В связи с этим вопрос о возможном влиянии кальция на биодоступность железа изучается длительное время как в экспериментах на животных, так и в исследованиях на людях.
Необходимо отметить, что клеточные механизмы всасывания, т. е. поступления ионов железа и кальция из просвета кишечника в ток крови через энтероциты кишечника, различны. Многочисленными работами было показано, что в этом процессе участвуют различные клеточные транспортеры (J. Hoenderop et al., 2005). Кроме того, имеются данные о том, что кальций уменьшает поступление в организм как гемового (L. Hallberg, 1991), так и негемового железа. Все вместе указывает на то, что кальций может влиять на биодоступность железа, оказывая ингибирующее влияние либо на транспорт его в желудочно-кишечном тракте, либо на связывание с рецепторами, расположенными на апикальной мембране энтероцитов.
В экспериментах на изолированный кишечной петле в условиях in vivo на крысах было показано уменьшение всасывания железа из раствора FeCl2, вводимого непосредственно в петлю при добавлении кальция. Причем эффект зависел от абсолютной концентрации кальция в двенадцатиперстной кишке, а не от молярного соотношения Ca/Fe (Barton et al., 1983). Изучение влияния на клеточный транспорт железа различных солей, содержащих кальций, показало, что наибольший ингибирующий эффект вызывает СаСО3, в то время как эффекты СаSO4 и Na2CO3 присутствуют, но в меньшей степени (Prather, 1992). Эта кальциевая соль, добавленная в количестве 500 мг, способна уменьшить всасывание негемового железа, содержащегося в пищевых продуктах на 32% в случае потребления пищи, не содержащей дополнительные ингибирующие вещества, и на 42% при потреблении продуктов в сочетании с яйцами, кофе и др. (Сook et al., 1991). СaCO3 уменьшает также всасывание железа при совместном использовании их в одной таблетке. В этом случае 300 мг кальциевой соли при совместном употреблении с 37 мг железа, присутствующего в виде FeSO4, уменьшает всасывание железа на 15% (Seligman et al., 1983; Cook et al., 1991).
На добровольцах провели исследования по усвояемости железа при совместном употреблении с кальцийсодержащими продуктами. Усвоение железа определяли радиоизотопным методом с использованием Fe55 и Fe59. Здоровые женщины (21 чел.) потребляли в течение 10 дней дополнительное количество молока и сыра (~ 930 мг кальция в день). Это привело к снижению абсорбции железа на 30–50% (Hallberg, 1995). На основании полученных данных авторы предполагают, что ингибирование всасывания железа происходит на этапе «просвет кишечника — энтероцит».
В исследованиях на людях также изучалось влияние искусственных минеральных добавок: сульфата железа, цитрата и фосфата кальция и др. Работа была проведена на 61 здоровом испытуемом. Для оценки всасывания использовался также двойной радиоизотопный метод. При употреблении цитрата кальция (600 мг) абсорбция железа снижалась на 49%, фосфата — на 62% (Cook et al., 1991). Интересно, что в этом исследовании эффект от применения кальциевых добавок отмечался только на фоне употребления пищи. Вероятно, конкуренция между катионами возникала при заполненном кишечнике. Теоретически возможно, что высокие концентрации кальция могут изменять реологические свойства пищевого комка в просвете верхней части тонкого кишечника (Conrad et al., 1993). На людях также изучалось различие во влиянии кальция на потребление гемового и негемового железа. Так, в исследованиях на 27 добровольцах с применением полного промывания кишечника для измерения степени усвоения железа при использовании кальциевых добавок (450 мг) было показано снижение абсорбции только гемового железа на 20%. В этой работе добавление кальция не влияло на абсорбцию негемового железа (Z. K. Roughead, 2005). В другом исследовании, проведенном на 44 мужчинах и 81 женщине, наблюдали снижение всасывания гемового железа из рациона при добавлении кальция в дозах от 40 до 300 мг. Максимальное снижение наблюдалось при дозе 300 мг и составило 74%. Дальнейшее увеличение содержания кальция до 600 мг не приводило к возрастанию ингибирования иона железа (L. Hallberg et al., 1991). Противоречивые результаты, получаемые в разных работах, связаны, по-видимому, со сложностью воспроизведения точности методических подходов, проводимых на людях.
Во всех приведенных выше исследованиях было показано в той или иной мере уменьшение абсорбции железа в желудочно-кишечном тракте на 20–60% при совместном употреблении с кальцийсодержащими продуктами в ходе однократного приема пищи или таблетированных препаратов. Характерно, что использованные дозы кальция не превышали дневную норму взрослого человека (во всех описанных случаях суммарное поступление кальция за сутки было меньше 1000 мг). Однако непосредственный механизм антагонистического влияния кальция на всасывание железа остается неясным.
Серия исследований, проведенных на добровольцах при длительном совместном приеме пищи, содержащей определенное количество железа и кальция, не позволила получить однозначного ответа о влиянии иона кальция на биодоступность железа, а главное — на уровень гемоглобина у этих испытуемых. Часто эффект выявлялся (ингибирование составляет 19%), но был статистически недостоверен (Reddy et al., 1997). По-видимому, длительные исследования на людях осложняются контролем над соблюдением диеты и составлением диеты для контрольной группы (S. R. Lynch, 2000).
Анализ литературы позволяет заключить, что экспериментальные исследования на животных и работы, проведенные на испытуемых, подтвердили, что ионы кальция способны уменьшать уровень всасывания железа. Степень выявления эффекта зависела от используемых методических подходов, которые в разных работах отличались друг от друга, и это затрудняет интерпретацию результатов. Однако возможность таких взаимодействий может быть наиболее актуальна и должна безусловно учитываться для людей, страдающих железодефицитными состояниями (анемии) или входящих в группу риска по этому состоянию (дети, беременные и т. д.). Для лечения и профилактики таких состояний необходимо увеличить потребление железа, как за счет соблюдения соответствующей диеты, так и с помощью минеральных добавок. Но следует помнить, что эффективность этих мер может значительно снижаться на фоне потребления диетического кальция или кальцийсодержащих витаминных комплексов. Ограничивать потребление кальция не желательно, поскольку во многих случаях (беременность, возраст 12–18 лет) существует повышенная потребность в обоих элементах. Выходом из ситуации может служить раздельное применение кальция и железа. Экспериментальные данные показали, что интервал между приемом кальция и железа даже в 4 ч исключает эффект ингибирования (A. Gleeprup et al., 1993). Помимо этого, во время приема препарата железа стоит воздержаться от употребления любых продуктов, содержащих кальций, т. е. требуется исключить весь спектр молочной продукции, а также зеленые части растений.
В данном случае удобно применять витаминно-минеральные комплексы, которые заранее предусматривают раздельное употребление железа и кальция. И это не единственное сочетание жизненно важных микронутриентов, проявляющих антагонистические свойства. Таким образом, грамотное разделение компонентов витаминно-минеральных комплексов по времени приема является необходимым условием эффективности их применения.
Литература
- Arredondo M., Martinez R., Nunez M. T. et al. Inhibition of iron and copper uptake by iron, copper and zinc. Biol. Res. 2006; 39: 95–102.
- Barton J. C., Conrad M. E., Parmley R. T. Calcium inhibition of inorganic iron absorption in rats. Gastroenterology. 1983; 84: 90–101.
- Conrad M. E., Umbreit J. N. A concise review: iron absorption- the mucin — mobilferrin — integrin parthway. A competitive parthway for metal absorption. Am J Hematology. 1993; 42: 67–73.
- Cook J. D. Adaption in iron metabolism. Am J Clin Nutrition. 1990; 42: 67–73.
- Cook J. D., Dassenko S. A., Whittaker P. Calcium supplementation: effect on iron absorption. Am J Clin Nutrition. 1991a; 53: 106–111.
- Donovan A., Brownile A., Zhou Y. et al. Positional cloning of zebrafish ferraportin identifies a conserved vertebrate iron exporter. Nature. 2000; 403: 776–781.
- Gleerup A., Rossander-Hulten L., Hallberg L. Duration of the inhibitory effect of calcium on non-haem iron absorption in man. Eur I Clin Nutr. 1993; 47: 875–879.
- Guinote, Fleming R., Silva R. et al. Using skin to assass iron accumulation in human metabolic disorders. Ion Beam Analysis. 2006; 249: 697–701.
- Hallenberg L., Brune M., Erlandsson M. et al. Calcium: effect of different amounts on nonheme- and heme-iron absorption in humans. Am J Clin Nutrition. 1991; 53: 112–119.
- Hoenderop J. Gj., Nilius B., Bindels R. J. M. Calcium absorption across epithelia. Physiol. Rev. 2005; 85: 373–422.
- Linder M. C. Nutrition and metabolism of the trace element. Nutritional Biochemestry and Metabolism. 1991: 151–198.
- Linder M. C., Moriya M., Whon A. et al. Vesicular transport of Fe and interaction with other metal ions in polarized Caco2 Cell monolayers. Biol. Res. 2006; 39: 143–156.
- Lynch S. R. The effect of calcium on iron absorption. Nutr. Res. Rev. 2000; 13: 141–158.
- Ma Y., Specian R. D., Yen K. Y. et al. The transcytosis of divalent metal transporter 1 and apo-transferrin during iron uptake in intesyinal epithelium. Am J Physiol. 2002; 283: 965–97.
- Prather Ta and Miller DD Calcium carbonate depresses iron bioavailability in rats more than calcium sulfate or sodium carbonate. J Nutrition. 1992; 122: 327–332.
- Reddy M. B. and Cook J. D. Effect of calcium on nonheme-ironabsorption from a complete diet. Am J Clin Nutrition. 1997; 65: 1820–1825.
- Roughead Z. K., Zito C. A., Hunt J. R. Inhibitory effects of dietary calcium on the initial uptake and subsequent retention of heme and nonheme iron in humans: comparisons using an intestinal lavage method. Am J Clin Nutr. 2005; 82(3): 589—597.
- Roy C. N. and Enns C. A. Iron homeostasis: New tales from the crypt. Blood. 2000; 96: 4020—4027.
- Seligman P. A., Caskey J. H., Frazier J. L. et al. Measuremants of iron absorption from prenatal multivitamin-mineral supplements. Obstetrics and Gynecology. 1983; 61: 356–362.
Н. А. Медведева, доктор биологических наук, профессор
МГУ, Москва